PNAS
Collapse of a fish population after exposure to a synthetic estrogen
http://www.pnas.org/cgi/content/full/104/21/8897
*Fisheries and Oceans Canada, Freshwater Institute, 501 University Crescent, Winnipeg, Manitoba, Canada R3T 2N6; and Molecular Indicators Research Branch, United States Environmental Protection Agency, 26 West Martin Luther King Drive, Cincinnati, OH 45268
Edited by Deborah Swackhamer, University of Minnesota, Minneapolis, MN, and accepted by the Editorial Board March 29, 2007 (received for review October 27, 2006)
Abstract
Municipal
wastewaters
are
a
complex mixture containing
estrogens
and
estrogen
mimics that
are
known to
affect
the reproductive health
of
wild fishes.
Male
fishes
downstream
of
some wastewater
outfalls
produce vitellogenin (VTG) (a
protein normally
synthesized by females
during oocyte maturation)
and
early-stage
eggs in their testes, and
this feminization
has
been attributed
to
the presence of
estrogenic
substances
such as
natural
estrogens
[estrone or 17![]() -estradiol
(E2)], the synthetic
estrogen
used in birth-control pills [17
-estradiol
(E2)], the synthetic
estrogen
used in birth-control pills [17![]() -ethynylestradiol
(EE2)], or weaker
estrogen
mimics such as
nonylphenol in the water.
Despite widespread
evidence that
male
fishes
are
being feminized, it is not known whether these low-level, chronic exposures
adversely
impact
the sustainability
of
wild populations.
We conducted a
7-year,
whole-lake
experiment at
the Experimental
Lakes
Area
(ELA)
in northwestern Ontario,
Canada,
and
showed that
chronic exposure
of
fathead
minnow (Pimephales
promelas)
to
low concentrations
(5–6 ng·L–1) of
the potent 17
-ethynylestradiol
(EE2)], or weaker
estrogen
mimics such as
nonylphenol in the water.
Despite widespread
evidence that
male
fishes
are
being feminized, it is not known whether these low-level, chronic exposures
adversely
impact
the sustainability
of
wild populations.
We conducted a
7-year,
whole-lake
experiment at
the Experimental
Lakes
Area
(ELA)
in northwestern Ontario,
Canada,
and
showed that
chronic exposure
of
fathead
minnow (Pimephales
promelas)
to
low concentrations
(5–6 ng·L–1) of
the potent 17![]() -ethynylestradiol
led to
feminization
of
males
through the production of
vitellogenin mRNA
and
protein, impacts
on gonadal
development as
evidenced by intersex in males
and
altered
oogenesis in females,
and,
ultimately,
a
near
extinction of
this species from the lake.
Our observations
demonstrate
that
the concentrations
of
estrogens
and
their mimics observed in freshwaters
can
impact
the sustainability
of
wild fish
populations.
-ethynylestradiol
led to
feminization
of
males
through the production of
vitellogenin mRNA
and
protein, impacts
on gonadal
development as
evidenced by intersex in males
and
altered
oogenesis in females,
and,
ultimately,
a
near
extinction of
this species from the lake.
Our observations
demonstrate
that
the concentrations
of
estrogens
and
their mimics observed in freshwaters
can
impact
the sustainability
of
wild fish
populations.
endocrine disrupters | fathead minnow | municipal wastewaters | population | level effects | whole-lake experiment
There is considerable evidence that fishes inhabiting waters that receive untreated municipal wastewaters or effluents from municipal wastewater treatment plants (MWTPs) are exposed to chemicals that affect reproductive endocrine function. Male fish downstream of some wastewater outfalls produce vitellogenin (VTG) mRNA and protein, associated with oocyte maturation in females, and early-stage eggs in their testes (1–3). This feminization has been linked to the presence of estrogenic substances such as the natural estrogen 17Despite growing documentation on the feminization of male fishes in waterways receiving municipal effluents, a critical question in the field of endocrine disruption research is whether these low-level, chronic exposures adversely impact wild populations (13). Although laboratory studies have shown decreased reproductive success of fish exposed to <1–5 ng·L–1 of EE2 (14, 15), it is unknown whether this response would be observed in wild populations and whether it would result in a subsequent decline in abundances. To assess the ecological risk posed by this class of compounds, we must understand population-level effects of estrogens and their mimics on aquatic organisms.
The fathead
minnow (Pimephales
promelas)
is a
common species in North America,
and
its range
extends from the southern United States
to
northern Canada
(16). It is an
important
food source for numerous game
fish
species, such as
lake
trout (Salvelinus
namaycush),
walleye
(Sander
vitreus), and
northern pike (Esox lucius). In the lakes
used for this study, fathead
minnow have
a
lifespan
of
![]() 4
years,
but few individuals
live past
2 years
of
age
(17). Asynchronous
spawning
starts
in early
summer and
extends for a
period of
4
years,
but few individuals
live past
2 years
of
age
(17). Asynchronous
spawning
starts
in early
summer and
extends for a
period of
![]() 2
months; multiple females
will typically
spawn
in the nest of
a
single male,
who will care
for developing eggs until hatching.
Sexual
maturity
occurs during the second year
of
life, so most fish
spawn
for only one season.
Therefore, 2 consecutive years
of
reproductive failure
cause
catastrophic
declines in abundance
(18, 19). Fathead
minnow have
been widely adopted
by the scientific community for toxicity
testing because
of
their widespread
distribution, ease
of
laboratory
culture, well characterized
biology and
life history,
and
the large
family
(Cyprinidae)
of
fishes
that
they represent (20–22).
It is a
freshwater
equivalent
of
the "miner's canary."
2
months; multiple females
will typically
spawn
in the nest of
a
single male,
who will care
for developing eggs until hatching.
Sexual
maturity
occurs during the second year
of
life, so most fish
spawn
for only one season.
Therefore, 2 consecutive years
of
reproductive failure
cause
catastrophic
declines in abundance
(18, 19). Fathead
minnow have
been widely adopted
by the scientific community for toxicity
testing because
of
their widespread
distribution, ease
of
laboratory
culture, well characterized
biology and
life history,
and
the large
family
(Cyprinidae)
of
fishes
that
they represent (20–22).
It is a
freshwater
equivalent
of
the "miner's canary."
We describe the results of a 7-year, whole-lake study at the Experimental Lakes Area (ELA) in northwestern Ontario, Canada, to assess the subcellular-level through population-level effects of the potent synthetic estrogen EE2 on fathead minnow. The concentrations of EE2 achieved in the experimental lake, Lake 260, during the 3 years of additions were within the range of those observed in untreated and treated municipal wastewaters (5, 6) and below the total E2 equivalents (with EE2 having an E2 equivalent of 0.19–1.9; ref. 23) measured in effluents and receiving waters (10–12).
Results and Discussion
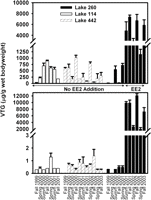
Measures of VTG mRNA and protein are used to assess exposure of fishes to estrogens and estrogen mimics, and elevated concentrations in males are indicative of the presence of these substances in effluents or surface waters (1, 3). We used these endpoints to assess the subcellular responses of fathead minnow from Lake 260 to EE2 additions. Seven weeks after the first estrogen additions to Lake 260 began in 2001, VTG concentrations in male and female fathead minnow were elevated when compared with preaddition or reference lake data (Fig. 1). Whole-body homogenates of males from Lake 260 had concentrations of VTG that were three orders of magnitude greater than reference samples, and this response was sustained in each of the 3 years of EE2 additions. Female fathead minnow also produced more VTG after exposure to EE2 when compared with reference samples (reference lake fish had 2.5% of the VTG concentrations observed in treated fish from Lake 260; Fig. 1), although their response was less dramatic than that observed for male fish. Liver VTG mRNA was measured in samples collected from the EE2-amended and reference lakes concurrently with those used for VTG protein analyses. Results from these mRNA analyses showed patterns similar to the VTG protein expression. Over the 3 years of dosing, male fathead minnow from Lake 260 had mean ± SD normalized liver VTG mRNA values ranging from 0.422 ± 0.685 to 1.22 ± 0.181. Liver VTG mRNA values for males from the two reference lakes were on average <0.1–1.6% of those for the EE2-exposed males. In addition, males from Lake 260 had VTG mRNA values (0.904 ± 0.437) that were more than an order of magnitude higher than female fish (0.045 ± 0.048) collected from the reference lakes during the same time periods. Females collected from Lake 260 in the spring and mid-summer of each year of EE2 additions had gene expression that ranged from 0.834 ± 0.879 to 0.378 ± 0.053, and this response was 82–152% of the values in females from the reference lakes. These among-lake differences were highest in the fall; for example, females collected in 2002 from Lake 260 exhibited levels of VTG mRNA expression (0.849 ± 0.793) that were more than an order of magnitude higher than those of reference females (0.045 ± 0.048). This trend was also observed in the accompanying protein data, demonstrating that EE2 amendments prolonged the expression of VTG mRNA and protein in males and females well past the breeding season.
|
|
Fig. 1. Mean ± SE (n = 4–7) VTG concentrations in whole-body homogenates of male (Lower) and female (Upper) fathead minnow captured in 1999–2003 from reference Lakes 114 and 442 and from Lake 260 before and during additions of 5–6 ng·L–1 of EE2 (low catches of fish in Lake 260 in 2004 and 2005 did not allow for these analyses in the latter 2 years of the study). |
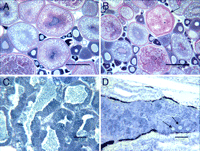
Wild fishes exposed to municipal wastewater effluents and fishes exposed to estrogens in laboratory studies have shown disruption in gonadal development as evidenced by the presence of both male and female gonadal tissues (i.e., intersex) and decreased gonadosomatic indices (GSIs) (2, 14, 15). In this study, testicular tissues of all of the male fathead minnow collected during the first spring after EE2 additions displayed delayed spermatogenesis, widespread fibrosis, and malformations of the tubules (see ref. 24). Testicular germinal tissue from all EE2-exposed males consisted primarily of spermatogonia instead of the spermatocytes that were normally observed at this time of year in the reference lakes and in Lake 260 before EE2 additions. In the spring of 2002, the GSI for males from Lake 260 averaged 0.40 ± 0.21% (n = 10). This mean was well below the GSI values of 1.39 ± 0.38% (n = 15) and 2.27 ± 0.41% (n = 10) recorded for males from reference Lakes 114 and 442, respectively. This arrested testicular development continued in 2003 and 2004, and four of nine males captured in the spring of 2003 had ova-testes with the presence of primary-stage oocytes (Fig. 2). It has been shown previously that male fish with intersex have reduced fertilization success because of decreased sperm production and mobility (25).
|
|
Fig. 2. Gonadal tissue sections from fathead minnow sampled in early May 2003. (A) Ovary from reference Lake 442 with small, dark, primary-stage oocytes situated between cortical alveolar-stage oocytes and large vitellogenic oocytes. (B) Ovary from EE2-amended Lake 260 demonstrating an atretic follicle (arrow). (C) Testis from reference Lake 442. (D) Testis from EE2-amended Lake 260 demonstrating intersex; arrows indicate primary-stage oocytes among the remnants of testicular tissue. (Scale bars: A, B, 300 µm; C, D, 80 µm.) |
Index trap netting [adults and young-of-the-year (YOY)] and minnow traps (adults) were used to examine changes in the fathead minnow population size and structure in the fall during each of the 7 years in both Lake 260 and Lake 442 (Fig. 3). Trap-net catch-per-unit-effort (CPUE) data showed that the mean abundance of this species varied considerably (9.6 ± 2.2 to 490 ± 68) in reference Lake 442 from 1999 to 2005 (Fig. 3A). In contrast, the CPUE of fathead minnow decreased consistently after EE2 additions to Lake 260 from 180 ± 48 and 50 ± 43 (before additions) to 0.1 ± 0.01 in the fifth year after additions began (Fig. 3B). Lake 260 and Lake 442 fathead minnow CPUE data derived from fall catches using minnow traps (data not shown) showed the same trends as the trap-net data and, overall, the two methods were highly comparable (Spearman rank correlation: t12 = 6.45, R = 0.88, P < 0.0001). Although fathead minnow populations typically fluctuate in size from year to year because of variable recruitment, year class failures are not observed in unmanipulated lakes (17, 27).
|
|
Fig. 3. Length frequency distributions of fathead minnow captured in trap nets in reference Lake 442 (A) and Lake 260 (B) (amended with 5–6 ng·L–1 of EE2 in 2001–2003) during the fall of 1999–2005. Distributions for each fall have been standardized to 100 trap-net days. Mean ± SE daily trap-net CPUE data for adults and juveniles for the fall catches are shown in the panels. |
The results from this whole-lake experiment demonstrate that continued inputs of natural and synthetic estrogens and estrogen mimics to the aquatic environment in municipal wastewaters could decrease the reproductive success and sustainability of fish populations. Chronic exposure of fathead minnow to a potent synthetic estrogen led to feminization of males through production of VTG mRNA and protein, continued production of VTG in females beyond the normal breeding season, impacts on gonadal development, as evidenced by intersex in males and altered oogenesis in females, and a near extinction of this species from the lake. Likely because of its short life-cycle, the fathead minnow was the first to show population collapse in this experiment. This response implies that short-lived fish species may generally be at greatest risk from exposure to estrogens and their mimics, but chronic exposure of longer-lived species to these substances may result in the loss of these populations as well.
Materials and Methods
The experimental system, Lake 260, is located in an undisturbed watershed at the Experimental Lakes Area, has a surface area of 34 ha and a maximum depth of 14 m, and contains naturally reproducing populations of lake trout, white sucker (Catostomus commersoni), pearl dace, and fathead minnow, as well as several other small-bodied fish species. We studied Lake 260 and two nearby reference systems, Lakes 114 and 442, for 2 years before EE2 additions, 3 years during the amendments to the experimental lake, and 2 years after the additions were discontinued. The physical, chemical, and biological characteristics of Lake 260 and the reference lakes are detailed in refs. 24 and 28.
We added EE2 (Schering AG, Berlin, Germany) three times weekly to Lake 260 starting in May 2001, after the lake had stratified, and additions were continuous during the open-water seasons of 2001–2003 for a period of 20–21 weeks. For each EE2 addition, the required mass [calculated by using its predetermined half-life in the water column (K.A.K., unpublished data), the volume of the epilimnion, and the mean concentration of EE2 measured the previous week] was dissolved in 30 ml of DIG-grade methanol (Caledon Laboratories, Georgetown, ON, Canada) and added to the propeller wash of the boat as it was driven in transects across the lake. Mean ± SD epilimnetic concentrations of EE2 were quantified weekly by using replicate samples collected at each of five sites around the lake. EE2 was extracted onto preconditioned C-18 columns (Supelco, Oakville, ON, Canada), eluted with 100% DIG methanol, and quantified along with the internal standard testosterone by using standard RIA techniques and GC-MS confirmation for 10% of the samples. Additional details on extraction methods and EE2 concentrations have been described in refs. 24 and 28. Seasonal mean ± SD EE2 concentrations in Lake 260 were 6.1 ± 2.8 (n = 189), 5.0 ± 1.8 (n = 169), and 4.8 ± 1.0 (n = 170) ng·L–1 in 2001–2003, respectively.
For VTG mRNA and protein analyses, fathead minnow were collected by using live-minnow traps in the spring and fall of 1999–2003 and in the summer of 2000 and 2001 from Lake 260 and two reference lakes (n = 4–10 individuals per sampling date for each analysis). Each fish was handled according to Canadian Animal Care protocols (approved by the Freshwater Institute Animal Care Committee) and individually euthanized in pH-buffered tricaine methanesulfonate (250 mg·L–1). For VTG analyses, whole fish were placed in sterile plastic bags, immediately frozen on dry ice, and kept at –90°C until analyzed. Whole-body homogenates were used to quantify the VTG by using the indirect competitive ELISA methods detailed previously (24). Reagent VTG from fathead minnow was obtained from the Core Biomarker Facility (University of Florida, Gainesville, FL). The primary antibody used for the analysis was a mouse anti-carp VTG monoclonal antibody (ND-2D3; Biosense Laboratories, Bergen, Norway) raised against VTG from common carp (Cyprinus carpio). Fish captured for VTG mRNA analysis were euthanized and dissected immediately after capture, and their livers were placed in RNAlater (Ambion, Austin, TX) for transport to the laboratory. Liver VTG mRNA was measured by using quantitative real-time PCR and normalized to 18S ribosomal gene expression, an invariant ribosomal gene, by using methods detailed previously (29).
Impacts on fathead minnow gonadal development in Lake 260 were best examined in the spring of each year when gonads were mature and not affected by the asynchronous spawning season. Fish were collected from Lake 260 and the two reference lakes each spring from 1999 to 2003 as described above; whole fish were euthanized, immersed in Bouin's solution, and processed by using standard histological techniques as described previously (30).
Index trap netting (adults and young-of-the-year) and minnow traps (adults only) were used to examine changes in the fathead minnow population size and structure in the fall during each of the 7 years in both Lake 260 and reference Lake 442. Each September, at least three trap nets were set in each lake and fished from 7 to 33 consecutive days; nets were emptied every 2–3 days. Lake 260 and Lake 442 CPUE data (not shown) were also derived from fall minnow trapping (30 traps set per lake for 10–11 consecutive days and emptied daily) and compared with CPUE data from fall trap netting (shown in Fig. 3).
Acknowledgements
We thank D. W. Schindler, G. T. Ankley, K. R. Munkittrick, R. Länge, and two anonymous reviewers for their helpful comments on this manuscript. Fish population sampling was conducted by S. M. Chalanchuk and L. S. Tate [Department of Fisheries and Oceans (DFO), Winnipeg, MB, Canada]. This research was supported by the American Chemistry Council Long Range Initiative Program (K.A.K.), the Fisheries and Oceans Canada (DFO) Environmental Sciences Strategic Research Fund (K.A.K., V.P.P., R.E.E., P.J.B., and K.H.M.), the Canadian Federal Toxic Substances Research Initiative (K.A.K., V.P.P., and R.E.E.), and the Canada Research Chair program (K.A.K). EE2 was supplied by Schering AG. VTG analyses were done by K. Wautier (DFO, Winnipeg). EE2 analyses were conducted by L. Vandenbyllaardt (DFO, Winnipeg) and K. Londry (University of Manitoba, Winnipeg, MB, Canada).
Footnotes
Abbreviations:
CPUE, catch-per-unit-effort;
E2, 17![]() -estradiol;
EE2, 17
-estradiol;
EE2, 17![]() -ethynylestradiol;
VTG, vitellogenin.
-ethynylestradiol;
VTG, vitellogenin.
![]() To
whom correspondence should be sent at
the present address:
Canadian
Rivers Institute and
Biology Department,
University of
New Brunswick, Saint
John, New Brunswick, Canada
E2E 4P1. E-mail:
kiddk@unbsj.ca
To
whom correspondence should be sent at
the present address:
Canadian
Rivers Institute and
Biology Department,
University of
New Brunswick, Saint
John, New Brunswick, Canada
E2E 4P1. E-mail:
kiddk@unbsj.ca
Freely available online through the PNAS open access option.
Author contributions: K.A.K., P.J.B., K.H.M., and V.P.P. designed research; K.A.K., P.J.B., K.H.M., V.P.P., R.E.E., and J.M.L. performed research; P.J.B., K.H.M., V.P.P., R.E.E., J.M.L., and R.W.F. analyzed data; and K.A.K., P.J.B., K.H.M., V.P.P., R.E.E., J.M.L., and R.W.F. wrote the paper.
The authors declare no conflict of interest.
This article is a PNAS Direct Submission. D.S. is a guest editor invited by the Editorial Board.
© 2007 by The National Academy of Sciences of the USA
References